أوبابينيا
 العصر: المياولينغي (الوليوي)، 505 مليون سنة | |
|---|---|
 |
|
| المرتبة التصنيفية | جنس |
| التصنيف العلمي | |
| فوق النطاق | حيويات |
| مملكة عليا | أبواكيات |
| مملكة | بعديات حقيقية |
| عويلم | ثنائيات التناظر |
| مملكة فرعية | أوليات الفم |
| شعبة عليا | انسلاخيات |
| شعبة | مفصليات الأرجل |
| طائفة | قشريات رهيبة |
| الاسم العلمي | |
| Opabinia | |
| تعديل مصدري - تعديل | |
أوبابينيا ريغاليس هي مجموعة جذعية منقرضة من مفصليات الأرجل عاشت في فترة المياولينغي (أو ما يعرف بالكمبري الأوسط سابقا) في مراسب طفل برجس (قبل 505 ملايين سنة) في بريتيش كولومبيا.[1]
الأوبابينيا حيوان رخو الجسم، يصل قياسها إلى 7 سم طولًا، وجذعها المجزأ به زوائد على الجانبين وذيل على شكل مروحة. يظهر على الرأس سمات غير عادية: خمس عيون، وفم تحت الرأس موجه للخلف، وخرطوم ذو مخالب ربما كان يستخدم لتوصيل الطعام إلى الفم. ربما عاشت الأوبابينيا في قاع البحر مستخدمة خرطومها للبحث عن الغذاء الصغير والطري.[2] وُصِف منها أقل من عشرين عينة جيدة؛ 3 عينات منها عثر عليها في طبقة الفيلوبود العظمى، إذ تشكل أقل من 0.1 ٪ من مجتمع العينات.[3] عندما كشف الفحص الشامل الأول للأوبابينيا عام 1975 عن سماتها غير العادية، أُعتُقِد حينَها أن لا علاقة لها بأي شعبة معروفة، [4] أو ربما لها قرابة بأحد أسلاف المفصليات والديدان الحلقية. [2] ومع ذلك، ظهرت دراسات لاحقة منذ أواخر تسعينات القرن الماضي تدعم باستمرار قرابتها منهم بصفتها عضوا من المفصليات القاعدية، بجانب أقاربها من رتبة شعاعيات الأسنان (مثل الأنومالوكاريس، وأقاربها) التي تربط بينها وبين هذه الشعبة صلة قرابة وثيقة، وكذلك فصيات الأرجل ذات الخياشيم (مثل كريجماشيلا و بامبديلوريون).[5] [6] [7] [8] [1] [9] [10]
كان هناك نقاش مستمر في السبعينيات حول ما إذا كانت الحيوانات متعددة الخلايا ظهرت فجأة خلال أوائل الكمبري، فيما يسمى بالانفجار الكمبري، أو أنها نشأت في وقت سابق ولكن دون ترك أي أحافير. في البداية، اعتبرت الأوبابينيا دليلاً قوياً على فرضية «الانفجار». [4] إلا أنه في وقت لاحق، أدى اكتشاف سلاسل كاملة من فصيات الأقدام الشبيهة بها، والتي يشابه بعضها المفصليات إلى حد كبير، بالإضافة إلى تطور فكرة المجموعات الجذعية، أدى ذلك لاقتراح أن العصر الكمبري المبكر كان وقتًا للتطور السريع نسبيًا ولكن يمكن فهمه بالعمليات التطورية العادية دون افتراض أي عمليات تطورية فريدة من نوعها.[11]
تاريخ الاكتشاف
عدلوجد تشارلز دوليتل والكوت في طَفل برجس تسعة أحافير شبه كاملة لأوبابينيا ريغاليس وعدد قليل مما صنفه بـ أوبابينيا؟ ميديا، ونشر وصفًا وتصنيفها لهم عام 1912.[12] الاسم العام للأوبابينيا مشتق من ممر أوبابين في بريتيش كولومبيا في كندا. [12] بين عامي 1966 و 1967، وجد هاري ب. ويتينغتون عينة أخرى جيدة، [13] وفي عام 1975 نشر وصفًا تفصيليًا مبنيًا على تشريح شامل للغاية لبعض العينات وصور هذه العينات بإضاءات مختلفة من عدة زوايا. تحليل ويتينغتون لم يغطي أوبابينيا؟ ميديا: عينات والكوت من هذا النوع لم يكن بالإمكان التعرف عليها في مجموعته. [2] في عام 1960، وصف علماء أحافير روس العينات التي عثروا عليها في منطقة نوريلسكي في سيبيريا وسموها بـ أوبابينيا نورليسيا، [14] ولكن هذه الأحافير حفظت بشكل سيئ، فلم يشعر ويتينغتون أنها قدمت معلومات كافية لتصنيفها على أنها أعضاء في جنس أوبابينيا . [2]
التواجد
عدلجميع عينات الأوبابينيا المعترف بها حتى الآن عثر عليها في «طبقة فيلوبود» الموجودة في طفل برجس، في جبال روكي الكندية في بريتيش كولومبيا. [2]
أبلغ بريجز ونيدن في عام 1997 أنهما عثرا في طَفل إيمو (خليج إيمو) في جنوب أستراليا، على عينة جديدة من الميوسكوليكس المحفوظة جيداً بجودة أفضل مقارنة بسابقاتها، مما أدى بهم لاستنتاج أنها قريبة من الأوبابينا [15] - رغم أن هذا التفسير قد شك فيه دزيك لاحقًا، مختتماً استنتاجه بأن الميوسكوليكس مجرد دودة حلقية.[16]
المورفولوجيا
عدل-
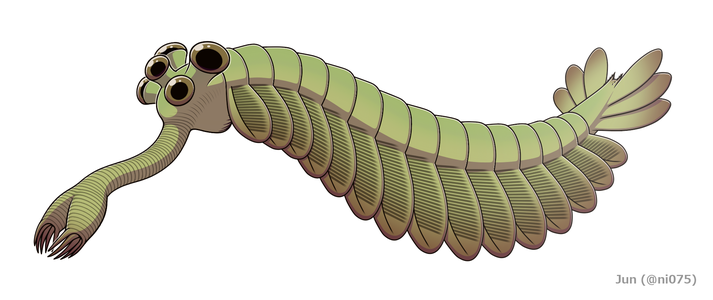 اعادة بناء
اعادة بناء -
 تقدير الحجم
تقدير الحجم


بدت الأوبابينيا غريبة جدًا لدرجة أن الجمهور ضحك في العرض الأول لتحليل ويتينغتون لها. [2] يتراوح طول أوبابينيا ريغاليس من الرأس (باستثناء خرطوم) إلى نهاية الذيل بين 4 سنتيمتر (1.6 بوصة) و 7 سنتيمتر (2.8 بوصة). [2] من أكثر السمات تميزًا في الأوبابينيا هو خرطومها المجوف، ويبلغ طوله الإجمالي حوالي ثلث الجسم ويتدلى من أسفل الرأس. خرطومها مرن، ومحزز مثل حزوز خراطيم المكنسة كهربائية، وينتهي خرطومها ببنية تشبه المخالب تحمل 5 أشواك في حافتها. يشير التناظر الثنائي والجانبي للخرطوم (بدلاً من التناظر الرأسي كما تصورها ويتينغتون عام 1975 [2] ) بأنه عبارة عن زوجٍ من الزوائد الأمامية المدمجة، مقارنة بتلك الموجدة لدى شعاعيات الأسنان، وفصيات الأقدام الخيشومية.[17] [5] [18] [19] يحمل الرأس خمس عيون معنقة (أي عيون لها عنق): اثنتان بالقرب من المقدمة وقريبة إلى حد ما من منتصف الرأس، متجهتان إلى الأعلى والأمام؛ وعينان أكبر بأعناق أطول بالقرب من الحواف الخلفية والخارجية للرأس، وتتجهان للأعلى والجانبين؛ وعين واحدة بين الزوج الأكبر من العيون المعنقة، وتتجه للأعلى. لقد اُفترض أن العيون كانت كلها مركبة، مثل العيون الجانبية الأخرى لمفصليات الأرجل [2] ولكن هذا التصور لا يدعمه أي دليل، [2] «خيالي إلى حد ما».[20] يقع الفم تحت الرأس، وخلف الخرطوم، ومتجه للخلف، بحيث يشكل الجهاز الهضمي انحناءًا على شكل حرف U في طريقه نحو مؤخرة الحيوان. ويتميز الخرطوم بالطول والمرونة بما يكفي للوصول إلى الفم. [2] الجزء الرئيسي من الجسم غالبا ما يكون عرضه حوالي 5 مليمتر (0.20 بوصة) ويحتوي على 15 جزءًا، يوجد على كل جزء أزواج من الرفارف (أو فصوص) متجهة للأسفل والخارج. تتداخل الرفارف بحيث يتم تغطية مقدمة كل رفرف منها بالحافة الخلفية للرفرف الأمامي. ينتهى الجسم بما يشبه قطعة مخروطية واحدة تحمل ثلاثة أزواج من شفرات الذيل المروحية المتداخلة، والتي تشير لأعلى وللخارج، لتشكل ذيلًا مثل مروحة مزدوجة على شكل حرف V. [2]
-
 تصورات مختلفة لبنية الرفرف مع الخياشيم، إذا يبين اللون الغامق الخياشيم، والفاتح الرفرف أو الفص.
تصورات مختلفة لبنية الرفرف مع الخياشيم، إذا يبين اللون الغامق الخياشيم، والفاتح الرفرف أو الفص. -
![مقطع عرضي للأوبابينيا، حسب بود ودالي (2011).[21]](//upload.wikimedia.org/wikipedia/commons/thumb/3/30/20210807_Opabinia_regalis_trunk_cross_section.png/420px-20210807_Opabinia_regalis_trunk_cross_section.png) مقطع عرضي للأوبابينيا، حسب بود ودالي (2011).[21]
مقطع عرضي للأوبابينيا، حسب بود ودالي (2011).[21]

![مقطع عرضي للأوبابينيا، حسب بود ودالي (2011).[21]](/wiki/%D9%85%D9%84%D9%81:20210807_Opabinia_regalis_trunk_cross_section.png)
تتباين وتختلف تفسيرات السمات الأخر لأحافير الأوبابينيا. نظرًا لأنها لم يكن لديها دروع متمعدنة حيوياً ولا حتى هياكل خارجية عضوية صلبة، كالموجودة لدى باقي مفصليات الأرجل، فإن أجسامها قد سُطحت أثناء طمرها وتحجرها، وتفاصيلها الأصغر أو الداخلية حُفظت كعلامات داخل الخطوط العريضة للأحفورة. [2] [5]
فسر ويتينغتون (1975) الخياشيم على أنها امتدادات متصلة ظهرياُ بقواعد كل الرفارف ما عدا الرفارف الأولى من كل جانب، واعتقد بأن الخياشيم المسطحة تحتها، عندها طبقات متداخلة من الجهة العلوية. [2] كشف بيرجستروم (1986) أن «الطبقات المتداخلة» عبارة عن صفوف من أنصال فردية، وفسر الرفارف بأنها جزء من أغطية ظهرية ( ترجات أو صفائح ظهرية) متموضعة على السطح العلوي من الجسم، ومزودة بشفرات مثبتة أسفل كل منها. [17] [8] اعتقد غراهام بود (1996) أن الشفرات الخيشومية متصلة على طول الحواف الأمامية من الجانب الظهري لكل الرفارف ما عدا الرفارف الأولى منها. و وجد أيضًا علامات داخل الحواف الأمامية من الرفارف، مفسراً إياها بأنها قنوات داخلية تربط الخياشيم بالجزء الداخلي من الجسم، مثلما فعل ويتينغتون عند تفسيره العلامة الظاهرة على طول الخرطوم بأنها قناة داخلية. [5] ومع ذلك، فسر تشانغ و بريغز (2007) أن جميع الرفارف لها تباعد خلفي حيث تتصل شفرات الخياشيم. [7] رفض بود و دالي (2011) إعادة البناء التي اقترحها تشانغ وبريغز، قائلا بأن الرفارف لها حواف خلفية كاملة كما اقترح في إعادة بناء سابقة. على الأغلب يتماشى تفسيرها مع إعادة البناء التي قام بها بود (1996) مع تعديلاتٍ على بعض التفاصيل (على سبيل المثال، يحتوي زوج الرفرف الأول على خياشيم أيضًا؛ ونقطة ربط الشفرات الخيشومية تقع في الخلف أكثر مما كان يعتقد في السابق). [8]


وجد ويتينغتون (1975) دليلاً لوجود ملامح شبه مثلثة على طول الجسم، وخلُص إلى أنها كانت هياكل داخلية، مرجحاً بأنها امتدادات جانبية للأمعاء (رُدب). [2] تشين وآخرون (1994) فسروها بأنها موجودة داخل الفصوص على طول جانبيها.[22] أما بود (1996) فيعتقد ان «المثلثات» شديدة الاتساع، أوسع من أن تُحتوى داخل جسم الأوبابينا النحيل، والمقطع العرضي بين بأنهم معلقين بشكل منفصل عن الفصوص وأخفض منها، وممتدة لأسفل الجسم. [5] وجد لاحقًا عينات حافظت على جليدة الساق الخارجية. لذلك فسر «المثلثات» بأنها أرجل قصيرة ولحميّة ومخروطية (فصيصات أرجل). كما وجد بقعًا متمعدنة صغيرة في أطراف بعضها، ففسرها بأنها مخالب. حسب هذا التصور وإعادة البناء، فإن الرفارف الحاملة للخياشيم وفصيصات أرجلها تتشابه مع التفرعات الخيشومية وتفرعات الأرجل الداخلية للأطراف ثنائية التفرغ عند المفصليات، والتي يمكن مشاهدتها لدى الماريلا وثلاثية الفصوص والقشريات. [5] قام تشانغ وبريغز (2007) بتحليل التركيب الكيميائي لـ«المثلثات»، وخلصا بأن لها نفس تركيبة القناة الهضمية، وبالتالي اتفقا مع ويتينغتون بأنهم جزء من الجهاز الهضمي. بدلا من ذلك، أعتبرا ترتيب الفصوص والخياشيم بأنها أحد الأشكال المبكرة لأطراف المفصليات قبل أن تتحول إلى أطراف ثناية التفرع. [7] ومع ذلك، فإن هذا التركيب الكيميائي المماثل لا يرتبط بالجهاز الهضمي فحسب؛ إذ يقترح بود ودالي (2011) أنه يمثل تمعدنًا يتشكل داخل تجاويف مملوءة بسائل داخل الجسم، وهو ما يتوافق مع فصيصات الأرجل المجوفة كما يظهر في الأحافير الفصيصية الواضحة. لقد وضحا أيضًا بأن رُتج أمعاء الأوبابينيا عبارة عن سلسلة من الغدد المعوية الدائرية المنفردة من «المثلثات». وعلى أنهم اتفقوا على عدم وجود مخالب طرفية، إلا أن وجود الفصيصات في الأوبابينيا يظل تفسيرًا معقولًا. [8]
أسلوب الحياة
عدلالطريقة التي طمرت بها حيوانات طفل برجس، عن طريق الانهيارات الطينية أو التيارات المحملة بالرواسب والتي كانت بمثابة عاصفة رملية آنذاك، تشير إلى أنها عاشت على قاع البحر. [2] ربما استخدمت الأوبابينيا خرطومها للبحث في الرواسب عن جزيئات الطعام وتمريرها إلى فمها. [2] ونظراً لعدم وجود أي علامة ولا أي دليل لوجود شيء يعمل كفكين عندها، فإنه من المفترض أن يكون طعامها صغيرًا ولينًا. [2] قد تزيد ردب القناة الهضمية من كفاءة هضم الطعام واستخلاص المغذيات.[23] يعتقد ويتينيغتون (1975) أن الأوبابينيا ليس لها أرجل، وأنها تزحف على فصوصها، ومن الممكن أيضًا أنها كانت تسبح ببطء من خلال خفق الفصوص، خاصةً إذا زامنت حركاتها لإنشاء حركة موجية متناسقة. [2] من ناحية أخرى، كان يعتقد أن جسمها لم يكن مرنًا بما يكفي ليسمح لها بتمويج جسمها كالأسماك. [2]
التصنيف
عدل| انسلاخيات |
| ||||||||||||||||||||||||||||||||||||





بالنظر إلى كيف اختلف علماء الأحافير في إعادة بناء الأوبابينيا وتصورهم عنها، فإنه من غير المستغرب أن يحتدم النقاش حول تصنيفها خلال القرن العشرين. [5] تشارلز دوليتل والكوت، الواصف الأصلي لها، اعتبرها قشريات عديمة الدرقة في عام 1912. [12] تبنى ج. إيفلين هاتشينسون هذه الفكرة عام 1930، إذ قدم أول إعادة بناء لأوبابينيا باعتبارها قشريات عديمة الدرقة تسبح مقلوبةً رأسًا على عقب.[24] قدم ألبرتو سيمونيتا إعادة بناء جديدة للأوبابينيا عام 1970 مختلفة كلياً عن إعادة بناء هاتشينسون، مع الكثير من خصائص المفصليات (مثل الهيكل الخارجي الظهري والأطراف المفصلية) التي تذكرنا باليوهويا واللينكويليا.[25] أعتقد ليف ستورمير، بعد العمل السابق الذي قام به بيرسي ريموند، أن الأوبابينيا تنتمي إلى ما يسمى بـ «الترايلوبودات» (مثل ثلاثية الفصوص وشبيهاتها). خلص ويتينغتون بعد تحليله الشامل عام 1975، إلى أن الأوبابينيا ليست من مفصليات الأرجل، إذ لم يعثر على أي دليل يشير لوجود أطراف مفصلية عندها، ولا يُعرف عن المفصليات أن لديها شيء مثل خرطوم الأوبابينيا المرن المملوء - ربما - بالسوائل. [2] وعلى أنه ترك تصنيف مستوى ما فوق العائلة مفتوحا، إلا أن جسمها الحلقي غير المتمفصل، ورفارفها الجانبية غير المألوفة ذات الخياشيم، أقنعه بأنها قد تمثل سلالة من أسلاف الحلقيات والمفصليات، [2] (العجلانيات العرفية والانسلاخيات، على التوالي) وهما شعبتان متمايزتان كان يُعتقد أنهما قريبتان آنذاك (تتحدان مع بعضهما حسب فرضية المتمفصلات).[26]
نشر ديريك بريجز وويتنغتون عام 1985 إعادة وصف رئيسية للأنومالوكاريس، المعثور عليها في طفل برجس.[27] بعد وقت قصير من ذلك، لاحظ عالم الأحافير السويدي يان بيرجستروم في عام 1986 التشابه بين الأنومالوكاريس والأوبابينيا، فاقترح أن الحيوانين كانا ذوي قربى، كما أنهما يشتركان في العديد من الخصائص (مثل الرفارف الجانبية، والشفرات الخيشومية، والعيون المعنقة والزوائد الأمامية المتخصصة). وصنفهما على أنهم مفصليات بدائية، على الرغم من أنه اعتبر أن المفصليات ليست شعبة واحدة. [17]
وجد جراهام بود في عام 1996 ما اعتبره دليلاً على وجود أرجل قصيرة غير مفصلية في الأوبابينيا. [5] دراسته للكيرجاماتشيلا المصنفة ضمن فصيات الأقدام ذوات الخياشيم من مرسب سيريوس باسيت الأحفوري، (مرسب أحفوري يعود عمره لحوالي 518 مليون سنة، أقدم من طفل برجس بـ 10 ملايين سنة)، أقنعه أن هذه العينة لها أرجل مشابهة.[28] [29] واعتبر أرجل هذين الجنسين مشابهة للغاية لأرجل الآييشيا من شعبة فصيات الأقدام الموجودة في طفل برجس و ومشابهة لأرجل حاملات المخالب (الديدان المخملية) الحديثة، التي تعتبر حاملة للعديد من سمات الأسلاف المشتركة مع أسلافها مع المفصليات. بعد فحص عدة مجموعات من السمات المشتركة بين هؤلاء والفصيات، رسم «إعادة بناء واسعة النطاق للمجموعة الجذعية الخاصة بالمفصليات» ، وبعبارة أخرى: إعادة بناء واسعة لمفصليات الأرجل والأعضاء الآخرين ممن اعتبرهم جذورهم التطوريون. [5] أحد الملامح البارزة لشجرة العائلة هذه هي أن بطيء الخطو الحديث (دب الماء) قد يكون أقرب قريب تطوري حي للأوبابينيا. [5] من ناحية أخرى، هيو وآخرون (1995، 2006) اقترحوا بدلاً من ذلك أن الأوبابينيا ما هي إلا عضوٌ من ديدان حلقيات الأعصاب غير العادية، مع ميزات تقاربية مع المفصليات. [30] [18]
على أن تشانغ وبريغز (2007) لا يتفقان مع تشخيص بود بأن «مثلثات» الأوبابينا كانت أرجلاً، إلا أن التشابه الذي رأوه بين ترتيب فصوص وخياشيم الأوبابينيا و الأطراف الثنائية للمفصليات قادهم إلى الاستنتاج بأن الأوبابينيا ذات صلة قرابة وثيقة بالمفصليات. في الواقع، لقد قدما شجرة عائلة تشبه إلى حد بعيد شجرة بود، باستثناء أن شجرتهم لم تذكر بطيئات الخطو (دب الماء). [7] بغض النظر عما اختلف العلماء فيه من تفسيرات مورفولوجية متباينة ومختلفة، إلا أن جميع اعادات الدراسة لها منذ الثمانينيات خلصت كلها أن التشابه بين الأوبابينيا ومفصليات الأرجل (كتشابه العيون المعنقة؛ و الظهر المجزء؛ والفم الخلفي؛ والزوائد المدمجة؛ وتفرعات الأطراف خيشومية الشكل) مهمة من الناحية التصنيفية. [17] [5] [7] [8] [1]
منذ عقد 2010، سقط الدعم عن علاقة القرب المقترحة بين الأوبابينيا وبطيء الخطو، مع حلقيات الأعصاب، وأصبح من المقبول أكثر ما يقترح من قرابة بين الأوبابينيا كمجموعة جذعية من المفصليات، مع شعاعيات الأسنان [9][10][1](وهي تفرع حيوي يندرج تحته الأنومالوكاريس وأقربائها[31]) ومفصليات الأرجل ذات الخياشيم، وقد كُشف عن هذا بواسطة التحاليل الوراثية [32][33][34][35][36][37][38][39][40][41][42][42] مع الكشوفات الجديدة مثل العثور على غدد معوية مشابه لتشريح المفصليات،[23][8] وكذلك الصنف الوسيط كيلينكسيا.[41]
الأهمية النظرية
عدل
بينت الأوبابينيا مدى ضآلة ما كان يُعرف عن الحيوانات رخوة الجسم، التي لا تترك أحافيرًا في العادة. [2] عندما وصفها ويتينغتون في منتصف السبعينيات، كان هناك بالفعل نقاش محتدم حول التطور المبكر للحيوانات. جادل بريستون كلاود بين عامي 1948 و 1968 بأن العملية التطورية كانت «متفجرة»، [43] وفي أوائل سبعينيات القرن الماضي، طور كل من نيلز إلدردج وستيفن جاي جولد نظريتهما عن التوازن النقطي، والتي تنظر إلى التطور على أنه فترات طويلة من شبه ركود يتخللها فترات قصيرة من التغيير السريع.[44] من ناحية أخرى وفي نفس الوقت تقريبا، جادل كل من وايت دورهام و مارتن غلايسنر بأن مملكة الحيوان كان لها تاريخ طويل مخفي في دهر الطلائع بسبب غياب الأحافير.[45] [46] خلص ويتنغتون (1975) إلى أن الأوبابينيا، وغيرها من الأصناف مثل الماريلا و اليوهويا ، لا يمكن إدراجها ضمن مجموعات الحيوانات الحديثة. كان هذا أحد الأسباب الرئيسية التي جعلت غولد يعتبر في كتابه الدائر موضوع عن طفل برجس، حياة رائعة، أن حياة الكمبري المبكرة كانت أكثر تنوعًا و«تجريبية» من أي مجموعة لاحقة من الحيوانات وأن الانفجار الكمبري كان حدثًا مثيرًا حقًا، ولربما كان مدفوعا بآليات تطورية غير عادية. [4] لقد اعتبر الأوبابينيا مهمة جدًا لفهم هذه الظاهرة لدرجة أنه أراد أن يطلق على كتابه «تكريما لأوبابينيا».[47]
ومع ذلك، سرعان ما تبع ذلك بعض الكشوفات والتحليلات، كاشفة عن حيوانات تشبهها مظهراً مثل الأنومالوكاريس من طفل برجس و الكيريجاماشيلا من سيروس باسيت. [17] [28] هناك حيوان آخر من طفل برجس، الآيشيا، قيل بأنها تشبه إلى حد بعيد حاملات المخالب الحديثة، [48] والتي تعتبر كأقارب للمفصليات.[49] عرّف علماء الأحافير مجموعة جديدة تسمى بفصيات الأرجل لكي تشمل أحافير «جميع-المفصليات» التي يعتقد أنها أقرباء لحاملات المخالب، وبطيء الخطو والمفصليات، ولكنها تفتقر للأطراف المفصلية. قُبلت هذه المجموعة لاحقاً على نطاق واسع باعتبارها درجة من درجات شبه العرق، مؤديةً لأصل شعبة «جميع-المفصليات» الموجودة.

- —= خطوط النسب
- = عقدة قاعدية
- = عقدة تاج
- = المجموعة الكلية
- = مجموعة تاج
- = مجموعة جذعية
بينما كانت الأوساط العلمية المختصة تتداول النقاش حول أحافير محددة مثل الأوبابينيا والأنومالوكاريس، في أواخر القرن العشرين، ظهر آنذاك مفهوم المجموعات الجذعية ليقدم فكرة «العمات» و «أبناء العمومة» التطوريون. مجموعة التاج هي مجموعة من الحيوانات الحية وثيقة الصلة بالإضافة إلى سلفهم المشترك الأخير وبالإضافة إلى جميع أحفادهم. تحتوي المجموعة الجذعية على فروع من أعضاء سلالة أقدم من آخر سلفٍ مشترك لمجموعة التاج؛ إنه مفهوم نسبي، على سبيل المثال، بطيئات الخطو هي حيوانات حية تشكل مجموعة تاج في حد ذاتها، لكن بود (1996) اعتبرها أيضًا مجموعة جذعية بالنسبة لمفصليات الأرجل. [5] [50] يمكن مشاهدة نسبية هذا المفهوم بالنظر إلى كائنات غريبة المظهر مثل الأوبابينيا بهذه الطريقة، نعم إن الانفجار الكمبري كان يبدو حدثاً غير عادي، إلا أنه يمكن فهمه من منظور العمليات التطورية العادية. [11]
انظر أيضًا
عدل- شعاعيات الأسنان - رتبة منقرضة من مفصليات الكمبري
اقرأ أيضا
عدل- Bergström, J. (1987). "The Cambrian Opabinia and Anomalocaris". Lethaia. ج. 20 ع. 2: 187–188. DOI:10.1111/j.1502-3931.1987.tb02037.x.
- Briggs, D. E. G.؛ Whittington, H. B. (1987). "The affinities of the Cambrian animals Anomalocaris and Opabinia". Lethaia. ج. 20 ع. 2: 185–186. DOI:10.1111/j.1502-3931.1987.tb02036.x.
روابط خارجية
عدل- "أوبابينيا ريغاليس" معرض أحافير طفل برجس، المتحف الافتراض الكندي، 2011
- صفحة الأوبابينيا في متحف سميثسونيان، مع صورة أحفورة من طفل برجس
مراجع
عدل- ^ ا ب ج د Briggs، Derek E. G. (19 أبريل 2015). "Extraordinary fossils reveal the nature of Cambrian life: a commentary on Whittington (1975) 'The enigmatic animal Opabinia regalis, Middle Cambrian, Burgess Shale, British Columbia'". Philosophical Transactions of the Royal Society B: Biological Sciences. ج. 370 ع. 1666: 20140313. DOI:10.1098/rstb.2014.0313. PMID:25750235.
{{استشهاد بدورية محكمة}}: الوسيط غير المعروف|PMCID=تم تجاهله يقترح استخدام|pmc=(مساعدة) - ^ ا ب ج د ه و ز ح ط ي يا يب يج يد يه يو يز يح يط ك كا كب كج كد Whittington, H. B. (يونيو 1975). "The enigmatic animal Opabinia regalis, Middle Cambrian Burgess Shale, British Columbia". Philosophical Transactions of the Royal Society B. ج. 271 ع. 910: 1–43 271. Bibcode:1975RSPTB.271....1W. DOI:10.1098/rstb.1975.0033. JSTOR:2417412. Free abstract at Whittington، H. B. (1975). "The Enigmatic Animal Opabinia regalis, Middle Cambrian, Burgess Shale, British Columbia". Philosophical Transactions of the Royal Society B. ج. 271 ع. 910: 1–43. Bibcode:1975RSPTB.271....1W. DOI:10.1098/rstb.1975.0033.
- ^ Caron، Jean-Bernard؛ Jackson، Donald A. (أكتوبر 2006). "Taphonomy of the Greater Phyllopod Bed community, Burgess Shale". PALAIOS. ج. 21 ع. 5: 451–65. Bibcode:2006Palai..21..451C. DOI:10.2110/palo.2003.P05-070R. JSTOR:20173022.
- ^ ا ب ج Gould, S. J. (1989). Wonderful Life. Hutchinson Radius. ص. 124–136 ff. ISBN:978-0-09-174271-3.
- ^ ا ب ج د ه و ز ح ط ي يا يب Budd, G. E. (1996). "The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group". Lethaia. ج. 29 ع. 1: 1–14. DOI:10.1111/j.1502-3931.1996.tb01831.x.
- ^ Budd, Graham E. (1998). "The morphology and phylogenetic significance of Kerygmachela kierkegaardi Budd (Buen Formation, Lower Cambrian, N Greenland)". Earth and Environmental Science Transactions of the Royal Society of Edinburgh (بالإنجليزية). 89 (4): 249–290. DOI:10.1017/S0263593300002418. ISSN:1473-7116. Archived from the original on 2021-11-04.
- ^ ا ب ج د ه Zhang, X.؛ Briggs, D. E. G. (2007). "The nature and significance of the appendages of Opabinia from the Middle Cambrian Burgess Shale". Lethaia. ج. 40 ع. 2: 161–173. DOI:10.1111/j.1502-3931.2007.00013.x.
- ^ ا ب ج د ه و Budd، G. E.؛ Daley، A. C. (2011). "The lobes and lobopods of Opabinia regalis from the middle Cambrian Burgess Shale". Lethaia. ج. 45: 83–95. DOI:10.1111/j.1502-3931.2011.00264.x.
- ^ ا ب Ortega-Hernández, Javier (2016). "Making sense of 'lower' and 'upper' stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848". Biological Reviews (بالإنجليزية). 91 (1): 255–273. DOI:10.1111/brv.12168. PMID:25528950. Archived from the original on 2021-10-07.
- ^ ا ب Ortega-Hernández, Javier; Janssen, Ralf; Budd, Graham E. (1 May 2017). "Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective". Arthropod Structure & Development (بالإنجليزية). 46 (3): 354–379. DOI:10.1016/j.asd.2016.10.011. ISSN:1467-8039. PMID:27989966. Archived from the original on 2021-11-29.
- ^ ا ب Budd, G. E. (2003). "The Cambrian Fossil Record and the Origin of the Phyla". Integrative and Comparative Biology. ج. 43 ع. 1: 157–165. DOI:10.1093/icb/43.1.157. PMID:21680420.
- ^ ا ب ج WALCOTT, C. D. 1912. Middle Cambrian Branchiopoda, Malacostraca, Trilobita and Merostomata. Smithsonian Miscellaneous Collections, 57: 145-228. نسخة محفوظة 2021-07-14 على موقع واي باك مشين.
- ^ Gould, S. J. (1990). Wonderful Life. London: Hutchinson Radius. ص. 77 and p. 189. ISBN:978-0-09-174271-3.
- ^ Miroshnikov, L. D. and Krawzov, A. G. (1960). Rare paleontological remains and traces of life in late Cambrian deposits of the northwestern Siberian platform. Palaeontology and biostratigraphy of the Soviet Arctic, 3, pp. 28–41.
- ^ Briggs, D. E. G.؛ Nedin, C. (يناير 1997). "The Taphonomy and Affinities of the Problematic Fossil Myoscolex from the Lower Cambrian Emu Bay Shale of South Australia". Journal of Paleontology. ج. 71 ع. 1: 22–32. DOI:10.1017/S0022336000038919. JSTOR:1306537.
- ^ Dzik, J. (2004). "Anatomy and relationships of the Early Cambrian worm Myoscolex". Zoologica Scripta. ج. 33 ع. 1: 57–69. DOI:10.1111/j.1463-6409.2004.00136.x.
- ^ ا ب ج د ه Bergström, J. (1986). "Opabinia and Anomalocaris, unique Cambrian arthropods". Lethaia. ج. 19 ع. 3: 241–246. DOI:10.1111/j.1502-3931.1986.tb00738.x.
- ^ ا ب Xianguang، Hou؛ Bergström، Jan (2006). "Dinocaridids – anomalous arthropods or arthropod-like worms?". في Jiayu، Rong؛ Zongjie، Fang؛ Zhanghe، Zhou؛ Renbin، Zhan؛ Xiangdong، Wang؛ Xunlai، Yuan (المحررون). Originations, Radiations and Biodiversity Changes – evidences from the Chinese fossil record. ص. 139–158. مؤرشف من الأصل في 2021-10-07.
- ^ Chipman، Ariel D. (18 ديسمبر 2015). "An embryological perspective on the early arthropod fossil record". BMC Evolutionary Biology. ج. 15 ع. 1: 285. DOI:10.1186/s12862-015-0566-z. ISSN:1471-2148. PMID:26678148. مؤرشف من الأصل في 2021-08-04.
{{استشهاد بدورية محكمة}}: الوسيط غير المعروف|PMCID=تم تجاهله يقترح استخدام|pmc=(مساعدة)صيانة الاستشهاد: دوي مجاني غير معلم (link) - ^ Paulus، H. F. (2000). "Phylogeny of the Myriapoda-Crustacea-Insecta: a new attempt using photoreceptor structure*". Journal of Zoological Systematics & Evolutionary Research. ج. 38 ع. 3: 189–208. DOI:10.1046/j.1439-0469.2000.383152.x.
- ^ Budd, Graham E.; Daley, Allison C. (2012). "The lobes and lobopods of Opabinia regalis from the middle Cambrian Burgess Shale". Lethaia (بالإنجليزية). 45 (1): 83–95. DOI:10.1111/j.1502-3931.2011.00264.x. ISSN:1502-3931. Archived from the original on 2021-05-25.
- ^ Chen, J-Y.؛ Ramsköld, L.؛ Gui-qing Zhou, G-Q. (مايو 1994). "Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators". Science. ج. 264 ع. 5163: 1304–1308. Bibcode:1994Sci...264.1304C. DOI:10.1126/science.264.5163.1304. PMID:17780848.
- ^ ا ب Vannier, Jean; Liu, Jianni; Lerosey-Aubril, Rudy; Vinther, Jakob; Daley, Allison C. (2 May 2014). "Sophisticated digestive systems in early arthropods". Nature Communications (بالإنجليزية). 5 (1): 3641. Bibcode:2014NatCo...5.3641V. DOI:10.1038/ncomms4641. ISSN:2041-1723. PMID:24785191. Archived from the original on 2021-10-05.
- ^ Hutchinson, George Evelyn (1930). Restudy of some Burgess shale fossils. Proceedings of the United States National Museum 78 (2854): 1–24. doi:10.5479/si.00963801.78-2854.1. نسخة محفوظة 2021-11-01 على موقع واي باك مشين.
- ^ Simonetta AM. 1970 Studies on non trilobite arthropods of the Burgess Shale (Middle Cambrian). Palaeontogr. Ital. 66, 35–45.
- ^ Edgecombe, Gregory D. (2009). "Palaeontological and Molecular Evidence Linking Arthropods, Onychophorans, and other Ecdysozoa". Evolution: Education and Outreach (بالإنجليزية). 2 (2): 178–190. DOI:10.1007/s12052-009-0118-3. ISSN:1936-6434.
- ^ Whittington، Harry Blackmore؛ Briggs، Derek Ernest Gilmor (14 مايو 1985). "The largest Cambrian animal, Anomalocaris, Burgess Shale, British-Columbia". Philosophical Transactions of the Royal Society of London. B, Biological Sciences. ج. 309 ع. 1141: 569–609. Bibcode:1985RSPTB.309..569W. DOI:10.1098/rstb.1985.0096. مؤرشف من الأصل في 2021-08-10.
- ^ ا ب Budd, G. E. (1993). "A Cambrian gilled lobopod from Greenland". Nature. ج. 364 ع. 6439: 709–711. Bibcode:1993Natur.364..709B. DOI:10.1038/364709a0.
- ^ Budd، G. E. (1997). "Stem Group Arthropods from the Lower Cambrian Sirius Passet Fauna of North Greenland". في Fortey؛ Thomas، R. H. (المحررون). Arthropod Relationships – Special Volume Series 55. Systematics Association.
- ^ Xian‐Guang، Hou؛ Bergström، Jan؛ Ahlberg، Per (1 سبتمبر 1995). "Anomalocaris and other large animals in the lower Cambrian Chengjiang fauna of southwest China". GFF. ج. 117 ع. 3: 163–183. DOI:10.1080/11035899509546213. ISSN:1103-5897. مؤرشف من الأصل في 2022-01-12.
- ^ Paleontological Society (United States) (0000 u). Journal of paleontology: a publication of the Society of Economic Paleontologists and Mineralogists and the Paleontological Society (بالإنجليزية). Bridgewater, MA [etc.: Society of Economic Paleontologists and Mineralogists. OCLC:819031091. Archived from the original on 10 يناير 2022.
{{استشهاد بكتاب}}: تحقق من التاريخ في:|تاريخ=(help) - ^ Legg، David A.؛ Sutton، Mark D.؛ Edgecombe، Gregory D.؛ Caron، Jean-Bernard (7 ديسمبر 2012). "Cambrian bivalved arthropod reveals origin of arthrodization". Proceedings. Biological Sciences. ج. 279 ع. 1748: 4699–4704. DOI:10.1098/rspb.2012.1958. ISSN:1471-2954. PMID:23055069. مؤرشف من الأصل في 2021-11-10.
{{استشهاد بدورية محكمة}}: الوسيط غير المعروف|PMCID=تم تجاهله يقترح استخدام|pmc=(مساعدة) - ^ Legg، David A. (2013). "Multi-Segmented Arthropods from the Middle Cambrian of British Columbia (Canada)". Journal of Paleontology. DOI:10.1666/12-112.1. مؤرشف من الأصل في 2021-12-26.
- ^ Lethaia: an international journal of palaeontology and stratigraphy (بالإنجليزية). London: Taylor & Francis. 0000 u. OCLC:66949846. Archived from the original on 3 نوفمبر 2021.
{{استشهاد بكتاب}}: تحقق من التاريخ في:|تاريخ=(help) - ^ Legg، David A.؛ Sutton، Mark D.؛ Edgecombe، Gregory D. (2013). "Arthropod fossil data increase congruence of morphological and molecular phylogenies". Nature Communications. ج. 4: 2485. DOI:10.1038/ncomms3485. ISSN:2041-1723. PMID:24077329. مؤرشف من الأصل في 2021-06-30.
- ^ Smith، Martin R.؛ Ortega‐Hernández، J. (2014). "Hallucigenia's onychophoran-like claws and the case for Tactopoda". Nature. DOI:10.1038/nature13576. مؤرشف من الأصل في 2022-01-08.
- ^ Roy، P.؛ Daley، A.؛ Briggs، D. (2015). "Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps". Nature. DOI:10.1038/nature14256. مؤرشف من الأصل في 2020-11-26.
- ^ Smith، Martin R.؛ Caron، Jean-Bernard (2015). "Hallucigenia's head and the pharyngeal armature of early ecdysozoans". Nature. DOI:10.1038/nature14573. مؤرشف من الأصل في 2021-01-21.
- ^ Caron، Jean-Bernard؛ Aria، Cédric (31 يناير 2017). "Cambrian suspension-feeding lobopodians and the early radiation of panarthropods". BMC evolutionary biology. ج. 17 ع. 1: 29. DOI:10.1186/s12862-016-0858-y. ISSN:1471-2148. PMID:28137244. مؤرشف من الأصل في 2021-10-29.
{{استشهاد بدورية محكمة}}: الوسيط غير المعروف|PMCID=تم تجاهله يقترح استخدام|pmc=(مساعدة)صيانة الاستشهاد: دوي مجاني غير معلم (link) - ^ Siveter، Derek J.؛ Briggs، Derek E. G.؛ Siveter، David J.؛ Sutton، Mark D.؛ Legg، David (2018-08). "A three-dimensionally preserved lobopodian from the Herefordshire (Silurian) Lagerstätte, UK". Royal Society Open Science. ج. 5 ع. 8: 172101. DOI:10.1098/rsos.172101. ISSN:2054-5703. PMID:30224988. مؤرشف من الأصل في 11 ديسمبر 2021.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة) والوسيط غير المعروف|PMCID=تم تجاهله يقترح استخدام|pmc=(مساعدة) - ^ ا ب Zeng، Han؛ Zhao، Fangchen؛ Niu، Kecheng؛ Zhu، Maoyan؛ Huang، Diying (2020). "An early Cambrian euarthropod with radiodont-like raptorial appendages". Nature. DOI:10.1038/s41586-020-2883-7. مؤرشف من الأصل في 2022-01-12.
- ^ ا ب "PALAEOBIOLOGY". PALAEOBIOLOGY. (بالإنجليزية). 0000 uuuu. ISSN:0094-8373. Archived from the original on 31 ديسمبر 2021.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(help) - ^ Cloud, P. E. (1948). "Some problems and patterns of evolution exemplified by fossil invertebrates". Evolution. ج. 2 ع. 4: 322–350. DOI:10.2307/2405523. JSTOR:2405523. PMID:18122310. and Cloud, P. E. (1968). "Pre-metazoan evolution and the origins of the Metazoa.". في Drake, E. T. (المحرر). Evolution and Environment. New Haven, Conn.: Yale University Press. ص. 1–72.
- ^ Eldredge, N. (1989). "APPENDIX: Punctuated Equilibria: An Alternative to Phyletic Gradualism". Time Frames: The Evolution of Punctuated Equilibria. ص. 193 et seq. DOI:10.1515/9781400860296.193. ISBN:9781400860296.
- ^ Jere H. (2004). Neoproterozoic-Cambrian Biological Revolutions: Presented as a Paleontological Society Short Course at the Annual Meeting of the Geological Society of America, Denver, Colorado, November 6, 2004 (بالإنجليزية). Yale University Reprographics & Imaging Service. Archived from the original on 2022-01-12.
- ^ Durham, J. W. (1971). "The fossil record and the origin of the Deuterostomata". Proceedings of the North American Paleontological Convention, Part H: 1104–1132. and Glaessner, M. F. (1972). "Precambrian palaeozoology". في Jones, J. B.؛ McGowran, B. (المحررون). Stratigraphic Problems of the Later Precambrian and Early Cambrian. University of Adelaide. ج. 1. ص. 43–52.
- ^ Knoll، A. H. (2004). "Cambrian Redux". The First Three Billion Years of Evolution on Earth. Princeton University Press. ص. 192. ISBN:978-0-691-12029-4. اطلع عليه بتاريخ 2009-04-22.
- ^ Robison, R. A. (يناير 1985). "Affinities of Aysheaia (Onychophora), with Description of a New Cambrian Species". Journal of Paleontology. ج. 59 ع. 1: 226–235. JSTOR:1304837.
- ^ Jacobs, D. K.؛ Wray, C. G.؛ Wedeen, C. J.؛ Kostriken, R.؛ DeSalle, R.؛ Staton, J. L.؛ Gates, R. D.؛ Lindberg, D. R. (2000). "Molluscan engrailed Expression, Serial Organization, and Shell Evolution". Evolution and Development. ج. 2 ع. 6: 340–347. DOI:10.1046/j.1525-142x.2000.00077.x. PMID:11256378.
- ^ ا ب Craske, A. J.؛ Jefferies, R. P. S. (1989). "A new mitrate from the Upper Ordovician of Norway, and a new approach to subdividing a plesion". Palaeontology. ج. 32: 69–99.