عين رأسيات الأرجل
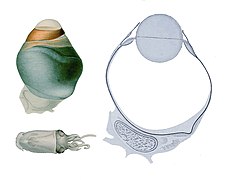
رأسيات الأرجل، باعتبارها مفترسات بحرية نشطة، تمتلك أعضاء حسية متخصصة للاستخدام في الظروف المائية.[1] لديها عين من نوع الكاميرا تتكون من قزحية وعدسة دائرية وتجويف زجاجي (جل العين) وخلايا صبغية وخلايا مستقبلة للضوء التي تحول الضوء من الشبكية الحساسة للضوء إلى إشارات عصبية تنتقل عبر العصب البصري إلى الدماغ. على مدى 140 عامًا، شكلت المقارنة بين عين رأسيات الأرجل من نوع الكاميرا وعين الفقاريات محورًا للبحث والدراسة كمثال على التطور المتقارب، حيث تطورت كلتا المجموعتين من الكائنات بشكل مستقل لتطوير صفة عين الكاميرا وتشترك كلتاهما في وظائف مشابهة. هناك جدل حول ما إذا كان هذا تطورًا متقاربًا حقًا أم تطورًا متوازيًا. على عكس عين الكاميرا للفقاريات، تتشكل عين رأسيات الأرجل كإنغماد من سطح الجسم (بدلاً من نتوءات من الدماغ)، وبالتالي تقع القرنية فوق العين بدلاً من أن تكون جزءًا هيكليًا من العين. على عكس عين الفقاريات، تُركز عين رأسيات الأرجل من خلال الحركة، مثل عدسة الكاميرا أو التلسكوب، بدلاً من تغيير الشكل كما تفعل العدسة في عين الإنسان. العين كروية تقريبًا، وكذلك العدسة، التي تكون داخلية بالكامل.[2][3][4]

تتطور أعين رأسيات الأرجل بطريقة تمر فيها محاور الشبكية خلف الشبكية، وبالتالي لا تحتاج العصب البصري إلى المرور عبر طبقة المستقبلات الضوئية للخروج من العين، وبالتالي لا تعاني من البقعة العمياء المركزية الفسيولوجية الطبيعية للفقاريات.[5]
يبدو أن بروتين الكريستالين المستخدم في العدسة قد تطور بشكل مستقل عن كريستالين الفقاريات، مما يشير إلى أصل متجانس للعدسة.[6]
تمتلك معظم رأسيات الأرجل أنظمة عضلية خارج العين معقدة تسمح بتحكم دقيق للغاية في تحديد موضع العينين بشكل عام. تمتلك الأخطبوطات استجابة ذاتية تحافظ على اتجاه بؤبؤ العينين بحيث تكون دائمًا أفقية.[1]
الضوء المستقطب
عدلعدة أنواع من رأسيات الأرجل، ولا سيما السبيدج والأخطبوط، وربما الحبار، لديها عيون قادرة على تمييز اتجاه الضوء المستقطب. ترجع هذه الحساسية إلى التنظيم العمودي لمستقبلات الضوء المجاورة. (لديها خلايا مستقبلة تسمى عيينة، مشابهة لتلك الموجودة في الرخويات الأخرى). على النقيض من ذلك، فإن عين الفقاريات عادة ما تكون غير حساسة للاختلافات في الاستقطاب لأن الأوبسينات في الخلية العصوية والمخروطية مرتبة بشكل شبه عشوائي. وبالتالي، فإن العين حساسة بالتساوي لأي اتجاه لمحور متجه الإلكترون للضوء. نظرًا لتنظيمها العمودي، فإن الأوبسينات في عيون رأسيات الأرجل لها أعلى امتصاص للضوء عندما تكون محاذية بشكل صحيح مع محور متجه الإلكترون للضوء، مما يسمح بالحساسية للاختلافات في الاستقطاب. لم تُثبت الوظيفة الدقيقة لهذه القدرة، ولكن يُفترض أنها تستخدم للكشف عن الفريسة والملاحة، وربما الاتصال بين رأسيات الأرجل التي تغير لونها.[7][8]
-
 عين باثيتيوثيس (جنس من السبيدج)
عين باثيتيوثيس (جنس من السبيدج) -
 عين الأخطبوط (الأخطبوط الشائع)
عين الأخطبوط (الأخطبوط الشائع) -
 عين السبيدج
عين السبيدج -
 عين الحبار
عين الحبار -
 عين البحار (البحار اللؤلؤي)
عين البحار (البحار اللؤلؤي)




.jpg)
الجدال التطوري
عدللا يزال هناك جدل قائم حول طبيعة تطور العين الكاميرا لدى رأسيات الأرجل والفقاريات: هل هو تطور مستقل من أصل واحد (تطور تقاربي)، أم تطور مشترك من سلف مشترك (تطور متوازي). وعلى الرغم من أن هذا الجدل قد حظي بدراسات واسعة، إلا أن الرأي السائد حاليًا يميل إلى أن تطور العين الكاميرا في كلا المجموعتين قد حدث بشكل مستقل من أصل واحد، أي تطور متقارب.
التطور المتوازي
عدلويقول الذين يصرون على أنه تطور متوازي إن هناك أدلة على وجود سلف مشترك يحتوي على المعلومات الجينية لتطور هذه العين. ويتضح ذلك من خلال احتواء جميع الكائنات ثنائية الجانب على الجين بروتين مربع مقترن (Pax6) الذي يعبر عن تطور العين.[9]
التطور التقاربي
عدلأولئك الذين يدعمون التطور التقاربي يذكرون أن هذا السلف المشترك كان سيسبق كل من رأسيات الأرجل والفقاريات بفارق كبير. كان السلف المشترك الذي يحمل تعبير العين من نوع الكاميرا موجودًا منذ حوالي 270 مليون سنة قبل تطور العين من نوع الكاميرا في رأسيات الأرجل وحوالي 110 إلى 260 مليون سنة قبل تطور العين من نوع الكاميرا في الفقاريات. مصدر آخر للدليل على ذلك هو الاختلافات في التعبير بسبب المتغيرات المستقلة لجين بروتين مربع مقترن سيكس التي ينشأ في كل من رأسيات الأرجل والفقاريات. تحتوي رأسيات الأرجل على خمسة متغيرات من هذا الجين في جينوماتها التي نشأت بشكل مستقل ولا تقوم بمشاركتها مع الفقاريات، على الرغم من أنها تسمح بتعبير جيني مشابه عند مقارنتها مع هذا الجين من الفقاريات.[10][11]
الأبحاث والاستخدام الطبي
عدلالاستخدام الطبي الرئيسي الناشئ في هذا المجال هو البحث في تطور العين والأمراض العينية. يجري حاليًا دراسات بحثية جديدة حول التعبير الجيني للعين باستخدام عيون رأسيات الأرجل نظرًا للأدلة على تطورها المتقارب مع العين البشرية الشبيهة. تحل هذه الدراسات محل دراسات ذبابة الفاكهة السابقة للتعبير الجيني أثناء تطور العين باعتبارها الأكثر دقة، على الرغم من أن دراسات ذبابة الفاكهة لا تزال الأكثر شيوعًا. يمنح الاستنتاج بأن العينين متشابهة مصداقية لمقارنتهما للاستخدام الطبي في المقام الأول، نظرًا لأن الصفة في كليهما قد تكون قد تشكلت من خلال الاصطفاء الطبيعي بواسطة ضغوط مماثلة في بيئات مماثلة؛ مما يعني وجود تعبير مماثل للأمراض العينية في عيني كلا الكائن الحي.[5]
من مزايا تجربة عين رأسيات الأرجل أنه يمكنها تجديد عيونها نظرًا لقدرتها على تجدد عملياتها التنموية، مما يسمح بمواصلة دراسات نفس رأسيات الأرجل بعد عينة تجريبية واحدة عند دراسة آثار المرض. كما يسمح ذلك بإجراء دراسة أكثر تعقيدًا تتعلق بكيفية الحفاظ على التجدد في الجينوم من رأسيات الأرجل وما إذا كان يمكن الحفاظ عليه إلى حد ما في الجينوم البشري إلى جانب الجينات التي تعبر عن الكاميرا العين.[5]
انظر أيضا
عدلالمراجع
عدل- ^ ا ب Budelmann BU. "Cephalopod sense organs, nerves and the brain: Adaptations for high performance and life style." Marine and Freshwater Behavior and Physiology. Vol 25, Issue 1-3, Page 13-33.
- ^ Serb, J.؛ Eernisse, D. (2008). "Charting Evolution's Trajectory: Using Molluscan Eye Diversity to Understand Parallel and Convergent Evolution". Evolution: Education & Outreach. ج. 1 ع. 4: 439–447. DOI:10.1007/s12052-008-0084-1.
- ^ Hanke، Frederike D.؛ Kelber، Almut (14 يناير 2020). "The Eye of the Common Octopus (Octopus vulgaris)". Frontiers in Physiology. ج. 10: 1637. DOI:10.3389/fphys.2019.01637. ISSN:1664-042X. PMC:6971404. PMID:32009987.
- ^ Yamamoto، M. (فبراير 1985). "Ontogeny of the visual system in the cuttlefish, Sepiella japonica. I. Morphological differentiation of the visual cell". The Journal of Comparative Neurology. ج. 232 ع. 3: 347–361. DOI:10.1002/cne.902320307. ISSN:0021-9967. PMID:2857734. S2CID:24458056.
- ^ ا ب ج Serb، Jeanne M. (2008). "Toward Developing Models to Study the Disease, Ecology, and Evolution of the Eye in Mollusca*" (PDF). American Malacological Bulletin. ج. 26 ع. 1–2: 3–18. DOI:10.4003/006.026.0202. S2CID:1557944. مؤرشف من الأصل (PDF) في 2014-12-18. اطلع عليه بتاريخ 2014-11-18.
- ^ SAMIR K. BRAHMA1 (1978). "Ontogeny of lens crystallins in marine cephalopods" (PDF). Embryol. Exp. Morph. ج. 46 ع. 1: 111–118. PMID:359745.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: أسماء عددية: قائمة المؤلفين (link) - ^ Mathger، L.M.؛ Shashar, N.؛ Hanlon, R.T. (2009). "Do cephalopods communicate using polarized light reflections from their skin?". Journal of Experimental Biology. ج. 212 ع. Pt 14: 2133–2140. DOI:10.1242/jeb.020800. PMID:19561202.
- ^ Shashar، N؛ Rutledge، P؛ Cronin، T (1996). "Polarization vision in cuttlefish in a concealed communication channel?". Journal of Experimental Biology. ج. 199 ع. 9: 2077–2084. DOI:10.1242/jeb.199.9.2077. PMID:9319987.
- ^ Gehring، W. J. (2004). "Historical perspective on the development and evolution of eyes and photoreceptors". The International Journal of Developmental Biology. ج. 48 ع. 8–9: 707–717. DOI:10.1387/ijdb.041900wg. PMID:15558463.
- ^ Fernald، Russell D. (29 سبتمبر 2006). "Casting a genetic light on the evolution of eyes". Science. ج. 313 ع. 5795: 1914–1918. Bibcode:2006Sci...313.1914F. DOI:10.1126/science.1127889. PMID:17008522. S2CID:84439732.
- ^ Yoshida، Masa-aki؛ Yura، Kei؛ Ogura، Atsushi (2014). "Cephalopod eye evolution was modulated by the acquisition of Pax-6 splicing variants". Scientific Reports. ج. 4 ع. 4256: 4256. Bibcode:2014NatSR...4E4256Y. DOI:10.1038/srep04256. PMC:3942700. PMID:24594543.